Реферат: Пресноводный полип гидра. Строение гидры пресноводной Гидра реагирует на раздражение
Энциклопедичный YouTube
- гидра длинностебельчатая (Hydra (Pelmatohydra) oligactis , синоним - Hydra fusca ) - крупная, с пучком очень длинных нитевидных щупалец, в 2-5 раз превышающих длину её тела. Эти гидры способны к очень интенсивному почкованию: на одной материнской особи порой можно встретить до 10-20 ещё не отпочковавшихся полипчиков.
- гидра обыкновенная (Hydra vulgaris , синоним - Hydra grisea ) - Щупальца в расслабленном состоянии значительно превышают длину тела - приблизительно вдвое длиннее тела, а само тело сужается ближе к подошве;
- гидра тонкая (Hydra circumcincta , синоним - Hydra attenuata ) - тело этой гидры имеет вид тонкой трубочки равномерной толщины. Щупальца в расслабленном состоянии не превышают длину тела, а если и превышают, то очень незначительно. Полипы мелкие, изредка достигают 15 мм. Ширина капсул голотрих изориз превышает половину их длины. Предпочитает жить поближе к дну. Почти всегда прикрепляется на сторону предметов, которая обращена ко дну водоёма.
- гидра зелёная () с короткими, но многочисленными щупальцами, травянистого зелёного цвета.
- Hydra oxycnida - щупальца в расслабленном состоянии не превышают длину тела, а если и превышают, то очень незначительно. Полипы крупные, достигают 28 мм. Ширина капсул голотрих изориз не превышает половины их длины.
1 / 5
✪ Гидра- подводная хищница.wmv
✪ Гидра пресноводная
✪ Пресноводный полип Гидра. Онлайн подготовка к ЕГЭ по Биологии.
✪ Создание Гидры (+ EEVEE), полный урок. Create a Hydra in Blender (+ EEVEE Demo)
Субтитры
Тело гидры цилиндрической формы, на переднем конце тела (на околоротовом конусе) расположен рот, окружённый венчиком из 5-12 щупалец. У некоторых видов тело разделено на туловище и стебелёк. На заднем конце тела (стебелька) расположена подошва, с её помощью гидра передвигается и прикрепляется к чему-либо. Гидра обладает радиальной (одноосно-гетеропольной) симметрией. Ось симметрии соединяет два полюса - оральный, на котором находится рот, и аборальный, на котором находится подошва. Через ось симметрии можно провести несколько плоскостей симметрии, разделяющих тело на две зеркально симметричных половины.
Тело гидры - мешок со стенкой из двух слоёв клеток (эктодермы и энтодермы), между которыми находится тонкий слой межклеточного вещества (мезоглея). Полость тела гидры - гастральная полость - образует выросты, заходящие внутрь щупалец . Хотя обычно считают, что у гидры есть только одно ведущее в гастральную полость отверстие (ротовое), на самом деле на подошве гидры имеется узкая аборальная пора. Через неё может выделяться жидкость из кишечной полости, а также пузырёк газа. При этом гидра вместе с пузырьком открепляется от субстрата и всплывает, удерживаясь вниз головой в толще воды. Таким способом она может расселяться по водоёму. Что касается ротового отверстия, то у не питающейся гидры оно фактически отсутствует - клетки эктодермы ротового конуса смыкаются и образуют плотные контакты, такие же, как и на других участках тела . Поэтому при питании гидре каждый раз приходится «прорывать» рот заново.
Клеточный состав тела
Эпителиально-мускульные клетки
Эпителиально-мускульные клетки эктодермы и энтодермы образуют основную массу тела гидры. У гидры около 20 000 эпителиально-мускульных клеток.
Клетки эктодермы имеют цилиндрическую форму эпителиальных частей и формируют однослойный покровный эпителий . К мезоглее прилегают сократимые отростки данных клеток, образующие продольную мускулатуру гидры.
Эпителиально-мускульные клетки энтодермы направлены эпителиальными частями в полость кишки и несут по 2-5 жгутиков, которые перемешивают пищу. Эти клетки могут образовывать ложноножки, с помощью которых захватывают частицы пищи. В клетках формируются пищеварительные вакуоли.
Эпителиально-мускульные клетки эктодермы и энтодермы представляют собой две независимые клеточные линии. В верхней трети туловища гидры они делятся митотически, а их потомки постепенно смещаются либо в сторону гипостома и щупалец, либо в сторону подошвы. По мере перемещения происходит дифференцировка клеток: так, клетки эктодермы на щупальцах дают клетки стрекательных батарей, а на подошве - железистые клетки, выделяющие слизь.
Железистые клетки энтодермы
Железистые клетки энтодермы выделяют в полость кишки пищеварительные ферменты, которые расщепляют пищу. Эти клетки образуются из интерстициальных клеток. У гидры около 5000 железистых клеток.
Интерстициальные клетки
Между эпителиально-мускульными клетками находятся группы мелких, округлых клеток, называемых промежуточными, или интерстициальными (i-клетки). У гидры их около 15 000. Это недифференцированные клетки. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных. Промежуточные клетки обладают всеми свойствами мультипотентных стволовых клеток. Доказано, что каждая промежуточная клетка потенциально способна дать как половые, так и соматические клетки. Стволовые промежуточные клетки не мигрируют, однако их дифференцирующиеся клетки-потомки способны к быстрым миграциям.
Нервные клетки и нервная система
Нервные клетки образуют в эктодерме примитивную диффузную нервную систему - рассеянное нервное сплетение (диффузный плексус). В энтодерме есть отдельные нервные клетки. Всего у гидры около 5000 нейронов . У гидры имеются сгущения диффузного плексуса на подошве, вокруг рта и на щупальцах. По новым данным, у гидры имеется околоротовое нервное кольцо, сходное с нервным кольцом, расположенным на крае зонтика у гидромедуз.
У гидры нет четкого деления на чувствительные, вставочные и моторные нейроны. Одна и та же клетка может воспринимать раздражение и передавать сигнал эпителиально-мускульным клеткам. Тем не менее, есть два основных типа нервных клеток - чувствительные и ганглиозные. Тела чувствительных клеток расположены поперек эпителиального пласта, они имеют неподвижный жгутик, окружённый воротничком из микроворсинок, который торчит во внешнюю среду и способен воспринимать раздражение. Ганглиозные клетки расположены в основании эпителиально-мускульных, их отростки не выходят во внешнюю среду. По морфологии большинство нейронов гидры - биполярные или мультиполярные.
В нервной системе гидры присутствуют как электрические, так и химические синапсы . Из нейромедиаторов у гидры обнаружены дофамин, серотонин, норадреналин, гамма-аминомасляная кислота, глютамат, глицин и многие нейропептиды (вазопрессин, вещество Р и др.).
Гидра - наиболее примитивное животное, в нервных клетках которого обнаружены чувствительные к свету белки опсины . Анализ гена опсина гидры позволяет предположить, что опсины гидры и человека имеют общее происхождение .
Стрекательные клетки
Стрекательные клетки образуются из промежуточных только в области туловища. Сначала промежуточная клетка делится 3-5 раз, образуя кластер (гнездо) из предшественников стрекательных клеток (книдобластов), соединённых цитоплазматическими мостиками. Затем начинается дифференцировка, в ходе которой мостики исчезают. Дифференцирующиеся книдоциты мигрируют в щупальца. Стрекательные клетки наиболее многочисленные из всех клеточных типов, их у гидры около 55 000.
Стрекательная клетка имеет стрекательную капсулу, заполненную ядовитым веществом. Внутрь капсулы ввёрнута стрекательная нить. На поверхности клетки находится чувствительный волосок, при его раздражении нить выбрасывается и поражает жертву. После выстреливания нити клетки погибают, а из промежуточных клеток образуются новые.
У гидры есть четыре типа стрекательных клеток - стенотелы (пенетранты), десмонемы (вольвенты), голотрихи изоризы (большие глютинанты) и атрихи изоризы (малые глютинанты). При охоте первыми выстреливают вольвенты. Их спиральные стрекательные нити опутывают выросты тела жертвы и обеспечивают её удержание. Под действием рывков жертвы и вызванной ими вибрации срабатывают имеющие более высокий порог раздражения пенетранты. Шипы, имеющиеся у основания их стрекательных нитей, заякориваются в теле добычи, а через полую стрекательную нить в её тело вводится яд.
Большое количество стрекательных клеток находится на щупальцах, где они образуют стрекательные батареи. Обычно в состав батареи входит одна крупная эпителиально-мускульная клетка, в которую погружены стрекательные клетки. В центре батареи находится крупная пенетранта, вокруг неё - более мелкие вольвенты и глютинанты. Книдоциты соединены десмосомами с мускульными волокнами эпителиально-мускульной клетки. Большие глютинанты (их стрекательная нить имеет шипы, но не имеет, как и у вольвент, отверстия на вершине), видимо, в основном используются для защиты. Малые глютинанты используются только при передвижении гидры для прочного прикрепления щупальцами к субстрату. Их выстреливание блокируется экстрактами из тканей жертв гидры.
Выстреливание пенетрант гидры было изучено с помощью сверхвысокоскоростной киносъёмки. Оказалось, что весь процесс выстреливания занимает около 3 мс. В его начальной фазе (до выворачивания шипов) скорость его достигает 2 м/c, а ускорение составляет около 40 000 (данные 1984 года ); видимо, это один из самых быстрых клеточных процессов, известных в природе. Первым видимым изменением (менее чем через 10 мкс после стимуляции) было увеличение объёма стрекательной капсулы примерно на 10 %, затем объём снижается почти до 50 % от исходного. В дальнейшем выяснилось, что и скорость, и ускорение при выстреливании нематоцист были сильно недооценены; по данным 2006 года , на ранней фазе выстреливания (выбрасывание шипов) скорость этого процесса составляет 9-18 м/с, а ускорение составляет от 1 000 000 до 5 400 000 g. Это позволяет нематоцисте массой около 1 нг развивать на кончиках шипов (диаметр которых составляет около 15 нм) давление порядка 7 гПа, что сравнимо с давлением пули на мишень и позволяет пробивать достаточно толстую кутикулу жертв.
Половые клетки и гаметогенез
Как и всем животным, гидрам свойственна оогамия . Большинство гидр раздельнополы, но встречаются гермафродитные линии гидр. И яйцеклетки, и сперматозоиды образуются из i-клеток. Считается, что это особые субпопуляции i-клеток, которые можно отличить по клеточным маркерам и которые в небольшом количестве присутствуют у гидр и в период бесполого размножения.
Дыхание и выделение
Дыхание и выделение продуктов обмена происходит через всю поверхность тела животного. Вероятно, в выделении некоторую роль играют вакуоли, которые есть в клетках гидры. Главная функция вакуолей, вероятно, осморегуляторная ; они выводят излишки воды, которые постоянно поступают в клетки гидры путём осмоса .
Раздражимость и рефлексы
Гидры имеют сетчатую нервную систему. Наличие нервной системы позволяет гидре осуществлять простые рефлексы . Гидра реагирует на механическое раздражение, температуру, освещённость , наличие в воде химических веществ и на ряд других факторов внешней среды.
Питание и пищеварение
Гидра питается мелкими беспозвоночными - дафниями и другими ветвистоусыми, циклопами , а также олигохетами-наидидами. Есть данные о потреблении гидрами коловраток и церкарий трематод . Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение). Непереваренные остатки пищи выбрасываются через рот.
Так как у гидры нет транспортной системы, а мезоглея (слой межклеточного вещества между экто- и энтодермой) достаточно плотная, возникает проблема транспорта питательных веществ к клеткам эктодермы. Эта проблема решается за счёт образования выростов клеток обоих слоёв, которые пересекают мезоглею и соединяются через щелевые контакты . Через них могут проходить мелкие органические молекулы (моносахариды, аминокислоты), что обеспечивает питание клеток эктодермы.
Размножение и развитие
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведёт самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады - половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры - сильно упрощённые споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм . Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5-1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление , в результате которого образуется целобластула. Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция . Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз . Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путём расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
Рост и регенерация
Миграция и обновление клеток
В норме у взрослой гидры клетки всех трёх клеточных линий интенсивно делятся в средней части тела и мигрируют к подошве, гипостому и кончикам щупалец. Там происходит гибель и слущивание клеток. Таким образом, все клетки тела гидры постоянно обновляются. При нормальном питании «избыток» делящихся клеток перемещается в почки, которые обычно образуются в нижней трети туловища.
Регенеративная способность
Гидра обладает очень высокой способностью к регенерации . При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность - рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва - на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/200 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Гидра может регенерировать из взвеси клеток, полученных путём мацерации (например, при протирании гидры через мельничный газ). В экспериментах показано, что для восстановления головного конца достаточно образования агрегата из примерно 300 эпителиально-мускульных клеток. Показано, что регенерация нормального организма возможна из клеток одного слоя (только эктодермы или только энтодермы).
Фрагменты разрезанного тела гидры сохраняют информацию об ориентации оси тела организма в структуре актинового цитоскелета : при регенерации ось восстанавливается, волокна направляют деление клеток. Изменение структуры актинового скелета может привести к нарушениям в регенерации (образованию нескольких осей тела) .
Опыты по изучению регенерации и модели регенерации
Местные виды
В водоёмах России и Украины наиболее часто встречаются следующие виды гидр (в настоящее время многие зоологи выделяют кроме рода Hydra ещё 2 рода - Pelmatohydra и Chlorohydra ):
Симбионты
У так называемых «зеленых» гидр Hydra (Chlorohydra) viridissima в клетках энтодермы живут эндосимбиотические водоросли рода Chlorella - зоохлореллы. На свету такие гидры могут длительное время (более четырёх месяцев) обходиться без пищи, в то время как искусственно лишённые симбионтов гидры без кормления погибают через два месяца. Зоохлореллы проникают в яйцеклетки и передаются потомству трансовариально . Другие виды гидр в лабораторных условиях иногда удается заразить зоохлореллами, однако устойчивого симбиоза при этом не возникает.
Именно с наблюдений за зелёными гидрами начал свои исследования А. Трамбле .
На гидр могут нападать мальки рыб, для которых ожоги стрекательных клеток, видимо, довольно чувствительны: схватив гидру, малёк обычно выплёвывает её и отказывается от дальнейших попыток съесть.
К питанию тканями гидр приспособлен ветвистоусый рачок из семейства хидорид Anchistropus emarginatus .
Тканями гидр могут также питаться турбеллярии микростомы , которые способны использовать непереваренные молодые стрекательные клетки гидр в качестве защитных клеток - клептокнид .
История открытия и изучения
Видимо, впервые описал гидру Антонио ван Левенгук . Подробно изучил питание, движение и бесполое размножение, а также регенерацию гидры
Тело гидры имеет вид продолговатого мешочка, стенки которого состоят из двух слоёв клеток — эктодермы и энтодермы .
Между ними лежит тонкая студенистая неклеточная прослойка — мезоглея , служащая опорой.

Эктодерма формирует покров тела животного и состоит из нескольких видов клеток: эпителиально-мускульные , промежуточные и стрекательные .
Самые многочисленные из них — эпителиально-мускульные.

Эктодерма
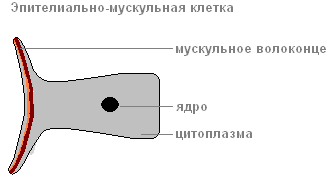
эпителиально-мускульная клетка
За счёт мускульных волоконец , лежащих в основании каждой клетки, тело гидры может сокращаться, удлиняться и изгибаться.
Между эпителиально-мускульными клетками находятся группы мелких, округлых, с большими ядрами и небольшим количеством цитоплазмы клеток, называемых промежуточными .
При повреждении тела гидры, они начинают усиленно расти и делиться. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных.
В эктодерме находятся стрекательные клетки , служащие для нападения и защиты. В основном они расположены на щупальцах гидры. Каждая стрекательная клетка содержит овальную капсулу, в которой свёрнута стрекательная нить.
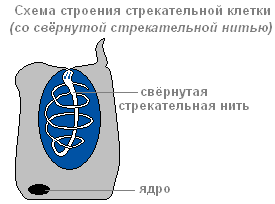
Строение стрекательной клетки со свернутой стрекательной нитью
Если добыча или враг прикоснётся к чувствительному волоску, который расположен снаружи стрекательной клетки, в ответ на раздражение стрекательная нить выбрасывается и вонзается в тело жертвы.
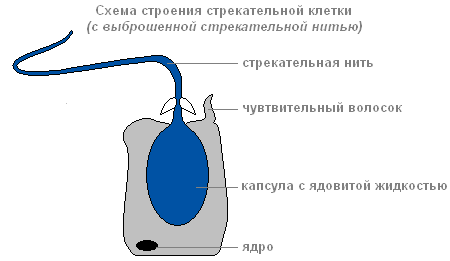
Строение стрекательной клетки с выброшенной стрекательной нитью
По каналу нити в организм жертвы попадает вещество, способное парализовать жертву.
Существует несколько типов стрекательных клеток. Нити одних пробивают кожные покровы животных и вводят в их тело яд. Нити других обвиваются вокруг добычи. Нити третьих — очень клейкие и прилипают к жертве. Обычно гидра «стреляет» несколькими стрекательными клетками. После выстрела стрекательная клетка погибает. Новые стрекательные клетки формируются из промежуточных .
Строение внутреннего слоя клеток
Энтодерма выстилает изнутри всю кишечную полость. В её состав входят пищеварительно-мускульные и железистые клетки.

Энтодерма
Пищеварительная система
Пищеварительно-мускульных клеток больше других. Мускульные волоконца их способны к сокращению. Когда они укорачиваются, тело гидры становится более тонким. Сложные движения (передвижение «кувырканием»), происходит за счёт сокращений мускульных волоконцев клеток эктодермы и энтодермы.
Каждая из пищеварительно-мускульных клеток энтодермы имеет 1-3 жгутика. Колеблющиеся жгутики создают ток воды, которым пищевые частички подгоняются к клеткам. Пищеварительно-мускульные клетки энтодермы способны образовывать ложноножки , захватывать и переваривать в пищеварительных вакуолях мелкие пищевые частицы.

Строение пищеварительно-мускульной клетки
Имеющие в энтодерме железистые клетки выделяют внутрь кишечной полости пищеварительный сок, который разжижает и частично переваривает пищу.

Строение желистой клетки
Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение ), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение ). Питательные вещества распределяются по всему телу гидры.
Когда в пищеварительной полости оказываются остатки жертвы, которые невозможно переварить, и отходы клеточного обмена, она сжимается и опорожняется.
Дыхание
Гидра дышит растворённым в воде кислородом. Органов дыхания у неё нет, и она поглощает кислород всей поверхностью тела.
Кровеносная система
Отсутствует.
Выделение
Выделение углекислого газа и других ненужных веществ, образующихся в процессе жизнедеятельности, осуществляется из клеток наружного слоя непосредственно в воду, а из клеток внутреннего слоя — в кишечную полость, затем наружу.
Нервная система
Под кожно-мускульными клетками располагаются клетки звездчатой формы. Это нервные клетки (1). Они соединяются между собой и образуют нервную сеть (2).

Нервная система и раздражимость гидры
Если дотронутся до гидры (2), то в нервных клетках возникает возбуждение (электрические импульсы), которое мгновенно распространяется по всей нервной сети (3) и вызывает сокращение кожно-мускульных клеток и всё тело гидры укорачивается (4). Ответная реакция организма гидры на такое раздражение — безусловный рефлекс .
Половые клетки
С приближением холодов осенью в эктодерме гидры из промежуточных клеток образуются половые клетки.
Различают два вида половых клеток: яйцевые, или женские половые клетки, и сперматозоиды, или мужские половые клетки.
Яйца находятся ближе к основанию гидры, сперматозоиды развиваются в бугорках, расположенных ближе к ротовому отверстию.
Яйцевая клетка гидры похожа на амёбу. Она снабжена ложноножками и быстро растет, поглощая соседние промежуточные клетки.

Строение яйцевой клетки гидры

Строение сперматозоида гидры
Сперматозоиды по внешнему виду напоминают жгутиковых простейших. Они покидают тело гидры и плавают с помощью длинного жгутика.
Оплодотворение. Размножение
Сперматозоид подплывает к гидре с яйцевой клеткой и проникает внутрь нее, причем ядра обеих половых клеток сливаются. После этого ложноножки втягиваются, клетка округляется, на ее поверхности выделяется толстая оболочка — образуется яйцо. Когда гидра погибает и разрушается, яйцо остается живым и падает на дно. С наступлением тёплой погоды живая клетка, находящаяся внутри защитной оболочки, начинает делиться, образующиеся клеточки располагаются в два слоя. Из них развивается маленькая гидра, которая выходит наружу через разрыв оболочки яйца. Таким образом, многоклеточное животное гидра в начале своей жизни состоит всего из одной клетки — яйца. Это говорит о том, что предки гидры были одноклеточными животными.

Бесполое размножение гидры
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведет самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады - половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры - сильно упрощенные споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм. Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5-1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление, в результате которого образуется целобластула. Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция. Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз. Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путем расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
Регенерация
Гидра обладает очень высокой способностью к регенерации. При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность - рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва - на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/100 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Передвижение
В спокойном состоянии щупальца вытягиваются на несколько сантиметров. Животное медленно водит ими из стороны в сторону, подстерегая добычу. При необходимости гидра может медленно передвигаться.
«Шагающий» способ передвижения
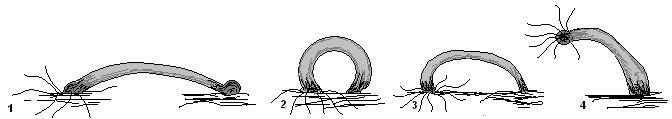
«Шагающий» способ передвижения гидры
Изогнув своё тело (1) и прикрепившись щупальцами к поверхности предмета (субстрата), гидра подтягивает к переднему концу тела подошву (2). Затем шагающее движение гидры повторяется (3,4).
«Кувыркающий» способ передвижения

«Кувыркающий» способ передвижения гидры
В другом случае она словно через голову кувыркается, поочерёдно прикрепляясь к предметам то щупальцами, то подошвой (1-5).
С.М. Никитина, И.А. Ваколюк (Калининградский государственный университет)
Функционирование гормонов как важнейших регуляторов и интеграторов метаболизма и разнообразнейших функций в организме невозможно без существования систем специфической рецепции сигнала и его трансформации в конечный полезный эффект, то есть без гормонкомпетентной системы . Иными словами, наличие реакции на организменном уровне на экзогенные соединения невозможно без наличия циторецепции к этим соединениям и соответственно без существования у этих животных эндогенных соединений, родственных тем, которыми мы воздействуем. Это не противоречит концепции универсальных блоков , когда основные молекулярные структуры в функциональных системах живых организмов обнаруживаются практически в полном наборе уже на самых ранних этапах эволюции, какие только доступны изучению, представлены ограниченным числом молекул и осуществляют одноименные элементарные функции не только у представителей одного царства, например в разных группах млекопитающих или даже в разных типах, но и у представителей различных царств, в том числе у многоклеточных и одноклеточных, у высших эукариот и прокариот.
Однако следует обратить внимание на то, что данные о составе и функциях соединений, выполняющих роль гормонов у позвоночных животных, у представителей таксонов достаточно низкого филогенетического уровня только начинают появляться . Из групп животных низкого филогенетического уровня - гидра как представитель кишечнополостных является наиболее примитивным организмом, обладающим настоящей нервной системой. Нейроны различаются морфологически, химически и, вероятно, функционально. Каждый из них содержит нейросекреторные гранулы . Установлено значительное разнообразие нейрональных фенотипов у гидры. В гипостоме имеются упорядоченно расположенные группы по 6-11 синаптически связанных клеток, которые можно рассматривать как доказательство наличия у гидр примитивных нервных ганглиев. Кроме обеспечения поведенческих реакций, нервная система гидр выполняет роль эндокринной регуляторной системы , обеспечивая контроль метаболизма, размножения, развития. У гидр существует дифференцировка нервных клеток по составу содержащихся в них нейропептидов ). Предполагают , что молекулы окситоцина, вазопрессина, половых стероидов и глюкокортикоидов являются универсальными. Они найдены и у представителей кишечнополостных. Головной и подошвенный активаторы (и ингибиторы) выделены из метаноловых экстрактов тела гидр. Головной активатор, выделенный из актиний, аналогичен по составу и свойствам нейропептиду, обнаруженному в гипоталамусе и кишечнике коровы, крысы, свиньи, человека и в крови последнего. Кроме того, было показано, что и у беспозвоночных и у позвоночных в обеспечении реакции клеток на нейрогормоны участвуют циклические нуклеотиды, то есть механизм действия этих веществ в двух филогенетически различных линиях един.
Целью данного исследования, учитывая вышеизложенное, мы избрали изучение комплексного влияния на пресноводную гидру экзогенных биологически активных (гормональных) соединений.
Материал и методы исследования
Животных для эксперимента собирали в июне-июле 1985-1992 гг. на стационаре (протока реки Немонин, поселок Матросово Полесского района). Адаптация к содержанию в лабораторных условиях - 10-14 суток. Объем материала: тип - Coelenterata; класс - Hydrozoa; вид - Hydra oligactis Pallas; количество - 840. Количество животных отражено в начале эксперимента и не учитывается прирост численности.
В работе были использованы водорастворимые гормональные соединения окситоцинового ряда, передней доли гипофиза с исходной активностью в 1 мл (ип) (гифотоцин - 5ЕД, питуитрин - 5ЕД, маммофизин - 3ЕД, префизон - 25ЕД, гонадотропин - 75ЕД) и стероид - преднизолон - 30 мг, которые у позвоночных обеспечивают трехзвенную эндокринную регуляцию, включающую гипоталамо-гипофизарный комплекс и эпителиальные железы.
В предварительных опытах были использованы концентрации препаратов от 0,00002 до 20 мл ип/л среды содержания животных.
Проводилось три группы исследования:
1-я - определение "+" или "-" реакции во всех принятых нами концентрациях;
2-я - определение диапазона концентраций, обеспечивающих работу в хроническом режиме разной продолжительности;
3-я - хронический эксперимент.
В эксперименте учитывалась активность почкования гидры. Полученные данные подвергались стандартной статистической обработке.
Результаты исследований
При определении "±" реакции гидр в широком диапазоне концентраций соединений были отобраны три (0,1 мл ип/л среды, 0,02 мл ип/л среды и 0,004 мл ип/л среды).
В контрольной группе гидр в течение пяти суток почкование оставалось на уровне 0,0- 0,4 почки /гидру (Ра). В среде минимальной концентрации префизона прирост был 2,2 особи/гидру, питуитрина - 1,9 особи/гидру (достоверность различий с контролем крайне высока - с уровнем значимости 0,01). В средних концентрациях хорошо себя проявили гифотоцин, маммофизин и префизон (1,8-1,9 особей/гидру). Преднизолон в минимальной, и особенно в средней концентрации, вызвал прирост численности 1,1-1,3 особи/гидру, что значительно превышает контроль.
В следующем эксперименте использованы только оптимальные концентрации гормональных соединений. Продолжительность эксперимента - 9 суток. К началу опыта по значению Ра контрольная и экспериментальная группы достоверно не различимы. Через девять суток эксперимента значения Ра достоверно отличалось в опытных группах и контроле с уровнем значимости 0,05 (табл. 1).
Таблица 1
Влияние гормональных препаратов на почкование гидры (Ра) и вероятность достоверности их различий (р)
| Среда | Ра | Изменение | р | ||
| 1 сутки | 9 сутки | Ра | 1 сутки | 9 сутки | |
| Контроль | 1,2±0,8 | 1,5±0,9 | 0,3±0,1 | - | - |
| Гонадотропин | 2,1±1,2 | 5,1±0,3 | 3,0±0,8 | 0,71 | 0,95 |
| Префизон | 1,1±0,7 | 4,9±2,0 | 3,8±1,3 | 0,13 | 0,97 |
| Гифотоцин | 1,8±0,8 | 6,1±2,2 | 4,3±1,4 | 0,58 | 0,99 |
| Питуитрин | 0,8±0,5 | 4,5±2,0 | 3,7±1,5 | 0,47 | 0,98 |
| Маммофизин | 1,1±0,3 | 5,3±2,0 | 4,2±1,7 | 0,15 | 0,99 |
| Преднизолон | 1,5±0,4 | 7,1±2,2 | 5,6±1,8 | 0,43 | 0,99 |
Как видно из таблицы, наибольшее значение Ра получено при содержании животных в преднизолоне. Все пептидные препараты дают приблизительно сходные значения Ра (в среднем 3,8±0,5). Однако и здесь есть разброс. Наилучший эффект (4,3±1,4) достигается при содержании животных в среде с очищенным экстрактом нейрогипофиза - гифотоцином. Близок к нему по степени воздействия маммофизин. В экспериментальных группах с питуитрином и префизоном значения величины Ра равны 3,7±1,5 и 3,8±1,3 соответственно. Наименьший эффект дает воздействие на гидр гонадотропином. Недостоверные различия в Ра возникают к концу первых суток после помещения гидр в растворы гормональных препаратов. На протяжении девяти суток эксперимента Ра в контроле не изменяется. Начиная с третьих суток Ра во всех экспериментальных группах существенно превышает Ра в контроле. Следует отметить постепенное достоверное увеличение этого показателя у экспериментальных групп к девятым суткам.
Для оценки статистической достоверности оказываемых воздействий сравнивались значения критерия F (отношение средних квадратов), полученные для каждого из двух факторов в отдельности (А - фактор длительности содержания; В - фактор воздействия) и для их взаимодействия (А+В), и табличные значения критерия для двух уровней значимости Р=0,05 и Р=0,01 (табл. 2).
Таблица 2
Результаты дисперсионного анализа влияния гормональных препаратов и длительности содержания на интенсивность бесполого размножения Hydra oligactis
| Фак- | Фактическое в группах | Табличое Р | ||||||
| торы | Питуитрин | Маммофизин | Гифотоцин | Гонадотропин | Префизон | Преднизолон | 0,05 | 0,01 |
| А | 3,44 | 1,40 | 2,27 | 2,17 | 3,62 | 1,30 | 1,92 | 2,50 |
| В | 8,37 | 4,04 | 8,09 | 4,73 | 8,26 | 12,70 | 4,00 | 7,08 |
| А+В | 1,12 | 0,96 | 0,56 | 0,37 | 1,07 | 1,03 | 1,92 | 2,50 |
Как видно из таблицы, Fфакт для фактора воздействия при уровне значимости 0,05 во всех экспериментальных группах больше Fтабл, а при уровне значимости 0,01 такая картина наблюдается в группах с питуитрином, гифотоцином, префизоном и преднизолоном, причем степень воздействия в группе с преднизолоном самая высокая, намного больше, чем в группах с питуитрином, гифотоцином и префизоном, имеющих сходную силу воздействия (значения Fфакт очень близки). Влияние взаимодействия факторов А и В во всех экспериментальных группах не является доказанным.
Для фактора А Fфакт меньше Fтабл (при обоих уровнях значимости) в группах с маммофизином и преднизолоном. В группах с гифотоцином и гонадотропином Fфакт больше Fтабл при Р=0,05, то есть влияние этого фактора не может считаться окончательно доказанным, в отличие от экспериментальных групп с питуитрином и префизоном, где Fфакт больше Fтабл и при Р=0,01 и при Р=0,05.
Все гормональные препараты, кроме гонадотропина, в той или иной степени задерживают начало бесполого размножения. Однако статистически достоверным это оказывается только в группе с префизоном (Р=0,01). Использованные в эксперименте гормональные препараты достоверно не влияют на продолжительность развития единственной почки, изменяют взаимное влияние первой и второй почек: питуитрин, маммофизин, префизон, гонадотропин - при наличии только сформированного головного отдела развивающихся почек; питуитрин, гонадотропин и преднизолон - при наличии хотя бы одного сформированного подошвенного отдела развивающихся почек.
Таким образом, можно считать установленной чувствительность гидр к широкому спектру гормональных соединений позвоночных и предположить, что экзогенные гормональные соединения включаются (как синергисты или антагонисты) в эндокринный регуляторный цикл, присущий самой гидре.
Список литературы
1. Перцева М.Н. Межмолекулярные основы развития гормонкомпетентности. Л.: Наука, 1989.
2. Богута К.К. Некоторые морфологические принципы формирования низкоорганизованных нервных систем в онто- и филогенезе // Успехи современной биологии. М.: Наука, 1986. Т. 101. Вып. 3.
3. Иванова-Казас А.А. Бесполое размножение животных. Л., 1971.
4. Наследов Г.А. Многовариантность осуществления элементарных функциональных задач и упрощение системы молекулярных взаимодействий как закономерность функциональной эволюции // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
5. Наточин Ю.В., Бройнлих Х. Использование методов токсикологии в изучении проблемы эволюции функций почки // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
6. Никитина С.М. Стероидные гомоны у беспозвоночных животных: Монография. Л.: Изд-во ЛГУ, 1987.
7. Афонькин С.Ю. Межклеточное самораспознование у простейших // Итоги науки и техники. М., 1991. Т. 9.
8. Проссер Л. Сравнительная физиология животных. М.: Мир, 1977. Т. 3.
9. Резников К.Ю., Назаревская Г.Д. Стратегия развития нервной системы в онто- и филогенезе. Гидра // Успехи современной биологии. М.: Наука, 1988. Т. 106. Вып.2 (5).
10. Шейман И.М., Балобанова Э.Ф., Пептидные гормоны беспозвоночных // Успехи современной биологии. М.: Наука, 1986. Т. 101. Вып. 2.
11. Этингоф Р.Н. Изучение молекулярной структуры нейрорецепторов. Методические подходы, эволюционные аспекты // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
12. Highnam K.C., Hill L. The comparative Endocrinology of the Invertebrates // Edward Arnold, 1
Движения гидры. Эпителиально-мускульные клетки эктодермы имеют во-локна, которые могут сокращаться. Если они сокращаются одновремен-но, укорачивается все тело гидры. Если сокращаются волокит в клетках с одной стороны, то в эту сторону гидра наклоняется. Благодаря работе этих волокон движутся щупальца гидры и перемещается все ее тело (рис. 13.4).
Реакции на раздражение гидры. Благодаря нервным клеткам, располо-женным в эктодерме, гидра воспринимает внешние раздражения: свет, прикосновения, некоторые химические вещества. Отростки этих клеток смыкаются между собой, образуя сетку. Так формируется самая простая по строению нервная система, называемая диффузной (рис. 13.5). Больше всего нервных клеток находится возле подошвы и на щупаль-цах. Проявлением работы нервной системы и эпителиально-мышечных клеток является безусловный рефлекс гидры — сгибание щупалец в от-вет на прикосновение.
| Рис. 13.4. Схема движении гидры |
 |
| Рис. 13.5. Нервная система гидры |
Во внешнем слое расположены также стрекательные клетки, содер-жащие капсулы со скрученной тоненькой трубочкой — стрекательной нитью. Наружу из клетки торчит чувствительный волосок. Достаточно его слегка коснуться, как нить выбрасывается из капсулы и вонзается в тело врага или добычи. К нему по стрекательной нити поступает яд, и животное погибает. Больше всего стрекательных клеток расположено в щупальцах.
Регенерация гидры. Маленькие округлые промежуточные клетки эктодер-мы способны превращаться в другие виды клеток. За счет их размноже-ния гидра быстро отстраивает поврежденную часть тела. Способность к регенерации этого животного поражает: когда гидру разделили на 200 частей, из каждой восстановилось целое животное!
Питание гидры. В эндодерме расположены железистые клетки и осна-щенные жгутиками пищеварительные клетки. Железистые клетки по-ставляют в кишечную полость вещества, называемые пищеваритель-ными соками. Эти вещества разрушают добычу, разлагая ее на микроскопические кусочки. С помощью жгутиков пищеварительные клетки подгоняют их к себе и захватывают, образуя псевдоподии. Внутреннюю полость гидры не случайно называют кишечной: в ней начинается переваривание пищи. Но окончательно пища расщепляет-ся в пищеварительных вакуолях пищеварительных клеток. Непе-реваренные остатки пищи удаляются наружу из кишечной полости через рот.
Выделение вредных веществ, образовавшихся в процессе жизнедея-тельности гидры, происходит через эктодерму в воду
Взаимодействие клеток. Среди клеток гидры лишь пищеваритель-ные переваривают пищу, но они обеспечивают питательными вещества-ми не только себя, но и все другие клетки. В свою очередь «соседи» создают наилучшие условия жизни для поставщиков питательных ве-ществ. Вспомните об охоте гидры — теперь вы сможете объяснить, как согласованная работа нервных, жалящих, эпителиально-мышечных и железистых клеток обеспечивает работой пищеварительные клетки. А эти клетки делятся с соседями результатами своего труда. Материал с сайта
Как размножается гидра? При бесполом размножении в результате деления промежуточных клеток образуется почка. Почка растет, на ней появляются щупальца, между ними прорывается рот. На противоположном конце формируется подошва. Маленькая гидра от-деляется от материнского организма, опускается на дно и начинает жить самостоятельно.
Размножается гидра и половым путем. Гидра — гермафродит: в од-них выступах ее эктодермы из промежуточных клеток формируются сперматозоиды, в других — яйцеклетки. Оставив тело гидры, спермато-зоиды по воде следуют к другим особям. Отыскав яйцеклетки, они оплодотворяют их. Образуется зигота, вокруг которой возникает плот-ная оболочка. Это оплодотворенное яйцо остается в теле гидры. Обычно половое размножение происходит осенью. Зимой взрослые гидры по-гибают, а яйца переживают зиму на дне водоема. Весной зигота начина-ет делиться, образуя два слоя клеток. Из них и развивается маленькая гидра.
На этой странице материал по темам:
Статеве розмноження губок
Раздражение и движение доклад биология
Особенности строения и процессов функционирования клеток тела гидры
Особенности процессов жизнедеятельности гидры
Сравнить строение стрекательной клетки гидры и кожицы листа крапивы.
Вопросы по этому материалу: